© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic"
На главную страницу сайта
Сайт наш чаще посещай - будет выше урожай (Лозунг времен Н.С.Хрущева)
|
© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic" |
|
Опубликовано: Гидробиологический журнал - 1998. - Т.34, № 5, стр. 94-104
УДК 574.58/282 4К
СТАТИСТИЧЕСКИЕ СВЯЗИ В ЭКОСИСТЕМЕ КУЙБЫШЕВСКОГО ВОДОХРАНИЛИЩА
В.В.Меншуткин, В.Н.Паутова, В.И.Номоконова, В А.Селезнев. И.И.Попченко, Т.Д.Зинченко, А.В.Иватин, Л.А.Выхристюк, М.М.Выхристюк, В.К. Шитиков, Т.И.Казанцева
Первое водохранилище на Волге — Иваньковское возникло в 1937 г., последнее — Чебоксарское в 1980 г. К настоящему времени накоплен большой фактический материал по экологии каждого из восьми крупных образовавшихся водохранилищ. Качество воды в их каскаде имеет важное значение для населения одного из крупнейших регионов России — Поволжья. Поэтому вполне понятен интерес к. обобщению полученных данных и pазработке на их основе экспертных оценок состояния экосистем волжских водохранилищ при разных вариантах водопользования и водопотреблении в регионе, антропогенных нагрузок и климатических флуктуаций.
В то же время обобщений на экосистемном уровне, сводящих в единое целое климатическую, экономическую, гидрологическую, гидрохимическую, санитарно-гигиеническую, гидробиологическую и рыбо-промысловую информацию за многолетний период в бассейне Волги, практически нет. Громадный фактический материал разбросан по сотням статей, тезисам докладов многочисленных конференций и отдельным монографиям Основное направление этих публикаций чаще всего заключатся в констатации фактов и достаточно скромных обобщениях на гидрологическом, гидрохимическом или гидробиологическом уровне.
Настоящая работа — одна из немногих попыток применения методов системного анализа экологии волжских водохранилищ. В ней рассмотрены результаты первого опыта эксплуатации банка данных по экологии Куйбышевского водохранилища — самого крупного в системе Волжского каскада и хорошо изученного. Здесь практически непрерывно с момента его образования наблюдали за гидрологическими и гидрохимическими условиями, динамикой численности и биомассы фито- и зоопланктона, за изменениями, происходящими в бактериопланктоне и донном населении водохранилища — бентосе [2, 12].
Эти материалы, а также данные наблюдений Гидрометеослужбы были оформлены в Институте экологии Волжского бассейна (ИЭВБ) РАН в виде компьютерного банка данных с использованием стандартной и широко распространенной системы DBASE [1], т.е. созданы предпосылки для использования системных методов в исследовании экосистемы водохранилища.
Материал и методика исследований. Наблюдения за состоянием экосистемы Куйбышевского водохранилища проводили с момента его образования (1957 г.) почти непрерывно до 1984 г. Станции отбора проб были постоянными и располагались по всей акватории, включая и речные участки по Каме — до Нижнекамской ГЭС и Волге — до Чебоксарской ГЭС. На каждой станции измеряли прозрачность воды, ее температуру у поверхности и у дна. определяли гидрохимические параметры, численность и биомассу бактериопланктона; в интегрированной пробе, охватывающей всю толщу воды, — состав фито- и зоопланктона, его численность, биомассу; отбирали пробы для изучения зообентоса. При обработке проб использовали стандартные методы [2]. Рейсы проводили в период открытой воды от трех до шести раз в год начиная с мая и кончая в октябре. При занесении этих материалов в банк данных использовали схему нумерации станций и их расположения, предложенную в литературе [12]. Кроме материалов многолетних наблюдений ИЭВБ РАН, банк данных содержал и информацию Гидрометеослужбы, характеризующую погодные и гидрологические условии в 1957—1984 гг.
Абиотические характеристики экологической системы в базе данных представлены солнечной радиацией (RAD) в районе г. Самары и пос. Вязовые, температурой приземного слоя воздуха (ТА, ТАМ] в районе г. Тольятти, температурой воды (ТЕМ), ее уровнем (LAV) в приплотинном плесе водохранилища и расходом (VVFL) через плотину ГЭС по материалам наблюдений Тольяттинской ГМО, а также температурой воды (TEMV) по данным экспедиций ИЭВБ РАН на каждой станции с подразделением на температуру поверхности воды (Ts) и придонного слоя (Тb).
Гидрохимические характеристики состоянии водоема включали концентрацию растворенного кислорода O2s и O2b — соответственно у поверхности и у дна, насыщение воды кислородом (О2 %). концентрации) растворенного углекислого газа (СО2), различных форм растворенного азота (NO2-, NO3-. и NH4+) и фосфатного фосфора (Р). Кроме того, использовали данные о концентрации водородных ионов (рН), кремния (SIL), растворенных форм железа (Fе), перманганатной окисляемости (РО) и цветности воды (COL). Бактериальное население водохранилища представлено обшей численностью бактерий (NBAC) и отдельно — численностью сапрофитных бактерий (NSAP), фитопланктонное сообщество — численностью диатомовых (NDIA, DIA), сине-зеленых (NCYA, CYA), пирофитовых (NPYR, PYR) и | зеленых (NCHL, CHL) водорослей. Зоопланктон характеризуется биомассами циклопоид (BCYC, CYC), каляноид (BCAL, CAL) и коловраток (BROT, ROT). Состояние бентоса водохранилища на каждой станции описывалось биомассой хирономид (RHIR, CHI), олигохет (BOLI, OLI), моллюсков (BMOL, MOL) и ракообразных (BCAR, CAR).
В исследование включены материалы наблюдений 1960—1984 гг. Всего массив использованных данных содержал более 50000 отдельных записей-лотов |8, 10, 11]. Поскольку в настоящем исследовании нас интересовали прежде всего связи между отдельными характеристиками экосистемы, то естественно было перейти к безразмерным величинам путем нормирования (1):
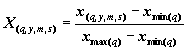
 (1)
(1)
где X(y, m, s) — нормированная характеристика, принимающая значения от 0 до 1; - x(y, m, s) — исходная характеристика экосистемы q; y - год; m - месяц; s - номер станции; xmin — минимальное значение этой характеристики по всему массиву данных. Переход к нормированным характеристикам экосистемы позволяет оценивать сходство или близость любых двух характеристик при помощи расстояния Хемминга [4]. Если суммирование абсолютных ззначений разности нормированных величин ведется ни всему множеству лет исследования {Y} и множеству станций водохранилища {S}, то результаты отображают близость сезонного хода , данных двух характеристик (2). Если суммирование ведется по всему множеству месяцев {М} и множеству станций {S}, то имеет место сходство многолетнего xода избранных характеристик экосистемы (3). Если суммирование охватывает все множество лет исследования {У}, все множество месяцев {М} и все множество характеристик системы, то результат свидетельствует о близости или похожести двух станций на акватории водохранилища (4):
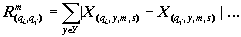 (2)
(2)
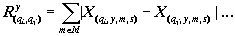 (3)
(3)
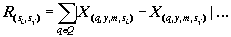 (4)
(4)
Здесь для экосистемы Куйбышевского водохранилища
Q - множество всех рассматриваемых характеристик:![]() Q = {RAD, ТАМ, LAV,.....BOLI, BHIR, BMOL};
Q = {RAD, ТАМ, LAV,.....BOLI, BHIR, BMOL};![]() (5)
(5)
У - множество лет исследования водохранилища:![]() У ={I960, 1961, .....1984}; (6)
У ={I960, 1961, .....1984}; (6)
М - множество месяцев:![]() М = {май, июнь, июль,.....октябрь};(7)
М = {май, июнь, июль,.....октябрь};(7)
S - множество станций:![]() S = {7,....20, 20а,...65, 66}.(8)
S = {7,....20, 20а,...65, 66}.(8)
После вычисления матрицы расстояний Хемминга между всеми характеристиками экосистемы были построены минимальные графы или “Вроцлавская таксономия” [6]. В терминах кластерною анализа эта процедура соответствует методу “ближнего соседа” [9].
Для понимания функционирования всей экосистемы было бы целесообразным одновременное рассмотрение всех характеристик со сдвигом во времени и пространстве. Однако для всего массива данных это оказалось задачей, находящейся за пределами возможности персонального компьютера, которым располагали авторы. Поэтому пришлось ограничиться только данными ст. 34 в Приплотинном плесе, как самыми полными. Были вычислены все коэффициенты линейной корреляции для 28 характеристик экосистемы в течение шести месяцев. Иными словами, получена корреляционная матрица размерностью 196´ 196, из которой отобраны только статистически значимые значения коэффициентов корреляции при доверительной вероятности 0,95.
Результаты исследовании и их обсуждение
Сезонные изменения структуры экосистемы. Все показатели по близости сезонных изменений группируются в отдельные плеяды (рис. 1). Так, сезонные изменения температуры воздуха над водохранилищем подобны изменениям температуры воды (ТЕМ), прихода солнечной радиации (RAD) и уровня водохранилища (LAV). Если в первых двух случаях связь имеет понятный причинно-следственный характер, то в третьем -- это типичная корреляционная зависимость: как и температура воздуха, уровень воды повышается весной при наполнении водохранилища и снижается к осени по мере его сработки. Такой же тип сезонной изменчивости имеют и связанные с температурой воды на станциях (TEMV) численность зеленых (NCHL), диатомовых (NDIA) и сине-зеленых (NCYA) водорослей, биомасса каляноид (BCAL), ракообразных бентоса (BCAR), моллюсков (BMOL), насыщение воды кислородом (О2 %), величина рН и концентрация растворенного в воде минерального фосфора (Р). То, что показатели температуры воды по данным Гидрометеослужбы (ТЕМ) и данным Института экологии Волжского бассейна (TEMV) близки, представляется очевидным. В основе близости сезонных изменений температуры воды, насыщения ее кислородом и численности основных групп водорослей — причинные связи, а вот похожесть сезонного хода биомассы моллюсков (BMOL), величины рН воды и ее прозрачности (TRAN) — факт нетривиальный и имеет, скорее всего, корреляционную основу.
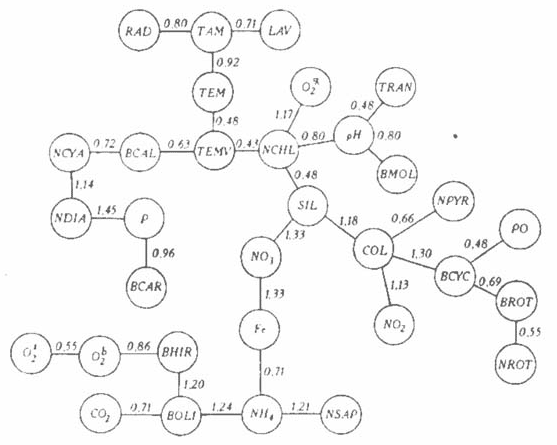
1. Минимальный граф связей сезонного хода характеристик экосистемы Куйбышевского водохранилища. Здесь и на рис. 2, 4: обозначения символов — в тексте. Здесь и на рис. 2: числа над линиями — расстояние Хемминга.
Подобными сезонными изменениями характеризуются также цветность коды (COL), содержание кремния (SIL), перманганатная окисляемость (РО|, биомасса циклопоид (BCYC) и коловраток (BROT), а также численность пирофитовых водорослей (NPYR) и содержание нитритного азота (NO2-). Эти показатели имеют весенний и осенний пики. Причины таких сезонных изменений в каждом случае различны. Например, цветность (COL) высока в весенних паводковых водах, она увеличивается и осенью при паводках и штормовых перемешиваниях водных масс. Кремний (SIL) поступает с паводковыми водами, затем потребляется интенсивно развивающимися диатомовыми водорослями; осенью при штормах его количество вновь повышается. В бимодальном характере кривой сезонной динамики циклопоид (BCYC) и коловраток (BROT) реализуются биологические особенности развивающихся в водохранилище видов. А нахождение в плеяде численности пирофитовых имеет, скорее всего, казуальный характер.
В другой группе показателей сходство сезонной динамики содержания аммонийного (NH4+) и нитратного (NO3-) азота, растворенного железа и численности сапрофитных бактерий (NSAP) может иметь причинную основу в отличие от похожести сезонных изменений биомассы хирономид (BHIR) в грунте и концентрации кислорода у поверхности воды (О2s) и у дна (О2b).
Не останавливаясь подробно на описании прочих связей сезонного хода характеристик экосистемы Куйбышевского водохранилища, отметим, что при анализе изученных данных (см. рис. 1) следует учитывать приближенность метода “ближнего соседа” и избегать соблазна поиска причинно-следственных связей там, где их, возможно, и нет или они весьма опосредованы. Смысл приведенных данных заключается вовсе не во вскрытии механизма сезонной динамики экологической системы, о в том, что для данного конкретного случая — экосистемы Куйбышевского водохранилища — найдены группы таких показателей, которые имеют сходный сезонный характер, что позволяет в перспективе качественно прогнозировать ход неизвестных величин по известным, в частности и в других водохранилищах Волги.
Межгодовые изменения. Соответствующий минимальный граф представлен на рис. 2. Отметим некоторые черты сходства графов сезонной и межгодовой динамики экосистемы, которые вовсе не очевидны априори. Например, близость рН и уровня насыщения воды кислородом (О2, %) (см. рис. 1 и 2) свидетельствует о том, что кислородный режим Куйбышевского водохранилища определяется процессом фотосинтеза фитопланктона, как, например, в олиготрофном Онежском озере. Другой случай совпадения сезонных и межгодовых тенденций — это близость функций цветности воды (COL) и концентрации нитритного (NO2-) иона (хотя прямой причинной связи тут, безусловно, нет) или вполне понятное сходство в сезонном и межгодовом аспектах динамики количества аммонийного азота (NH4+) и численности сапрофитов (NSAP).
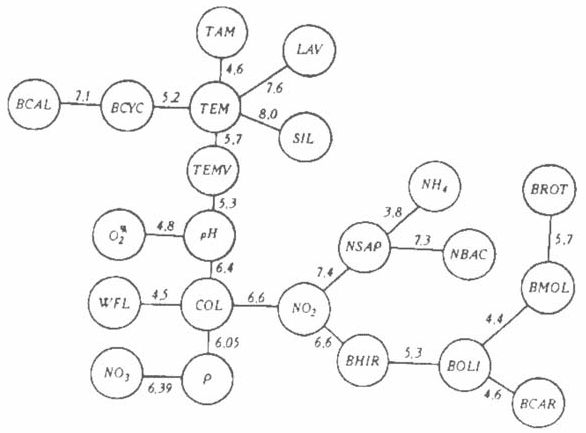
2. Минимальный граф связей межгодового хода характеристик экосистемы Куйбышевского водохранилища.
Характерно, что в межгодовом аспекте в противоположность сезонному все элементы бентоса (BHIR, BOLI, BMOL и BCAR) составляют единую группу со сходной динамикой. В прогнозных целях — это важный результат выполненного исследования, который может быть использован, например, при изучении антропогенных изменений экосистем и других водохранилищ, сходных с Куйбышевским по морфометрии и гидрологии. Что касается зоопланктона, то межгодовые флуктуации близки у представителей рачкового планктона (BCAL, BCYC).
Коловратки (BROT) имеют совершенно отличную от рачкового зоопланктона межгодовую, но сходную сезонную динамику биомассы. Это заставляет полагать, что в ряде случаев различаются факторы, определяющие сезонную и межгодовую изменчивость численности одних и тех же групп зоопланктона.
Последовательно связаны (см. рис. 2) температура воздуха (ТАМ), воды (ТЕМ, TEMV), рН, цветность (COL) и содержание минерального фосфора (Р). Расход воды в Волге (WFL) сказывается на среднегодовых значениях концентрации нитратного азота (NO3-), на биомассе бактерий (NSAP), эообентоса (BOLI, BCAR. BMOL, BROT), зоопланктона (BROT). Это также имеет важное прогностическое значение и совпадает с известным уже фактом повышения содержания биогенных элементов в многоводные годы вследствие интенсивного их выноса в водоем с площади водосбора. Интересно, что развитие каляноид (BCAZ) и циклопов (BCYC) больше связано с изменениями температуры воды (ТЕМ).
Среднегодовые значения численности отдельных групп фитопланктона оказались очень неустойчивыми и малоинформативными характеристиками — может быть, из-за меньшего периода наблюдений. Их пришлось исключить из рассмотрения в межгодовом аспекте. Необходимо отметить, что в использованном массиве данных имелись и другие досадные пропуски наблюдений. Регулярные исследования фитопланктона начались только в 1975 г., в данных по гидрохимии пропущены 1974 и 1980 гг. и т.п. Это не могло не сказаться на характере полученных зависимостей. Так, имеющийся материал не позволил вычислить характеристики экосистемы в данном году по сведениям за прошлые годы. Это имело бы важное прогностическое значение.
Пространственная структура. Для ее изучения были вычислены расстояния Хемминга между всеми станциями, на которых производились измерения. Граф минимальных расстояний между станциями приведен на рис. 3. Пожалуй, самым удивительным свойством этого графа является то, что он в общих чертах повторяет конфигурацию акватории водохранилища: последовательно связаны практически все станции, расположенные по руслу Волги. Этого совершенно нельзя было заметить по исходным данным. Следует обратить внимание на то, что камские воды (ст. 45) прослеживаются по совокупности всех характеристик до Ульяновского плеса (ст. 56). Это заставляет задуматься о сложности процессов смешения камских и волжских вод — вернее, даже не самих вод, а формирования экологических систем, с ними связанных. По русловым станциям связь прерывается между станциями 56 и 65, расположенными соответственно выше и ниже обширного мелководного Черемшанского залива. Очевидно, что влияние этого залива, загрязняемого отходами Сусканского рыбного хозяйства и предприятий г. Димитровграда (связанных в том числе и с атомной промышленностью), на жизнь водохранилища значительнее, чем представлялось до сих пор.
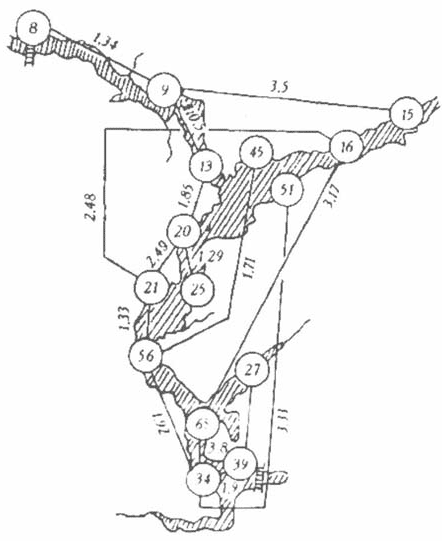
3. Расстояния Хемминга между станциями наблюдения на Куйбышевском водохранилище. Нумерация станций дана по [12].
Характерно, что графы, подобные изображенному на рис. 3, но построенные только по абиотическим характеристикам экосистемы и только по биотическим, оказались очень похожими на него и отличались только деталями. Рисунок может быть использован при обсуждении такой проблемы, как организация экологического мониторинга состояния водохранилища, в частности для определения станций проведения наблюдений. Если критерием взять принцип сходства, то на роль основной станции мониторинга может претендовать ст. 34, имеющая наибольшее количество связей с другими станциями по акватории водохранилища.
Функционирование экосистемы водохранилища. Корреляционные связи, рассчитанные со сдвигом пo времени по данным наблюдений на ст. 34, представлены на рис. 4. Последний имеет несколько упрощенный вид: опущены связи с отрицательными коэффициентами и некоторые характеристики для того, чтобы сделать его обозримым.
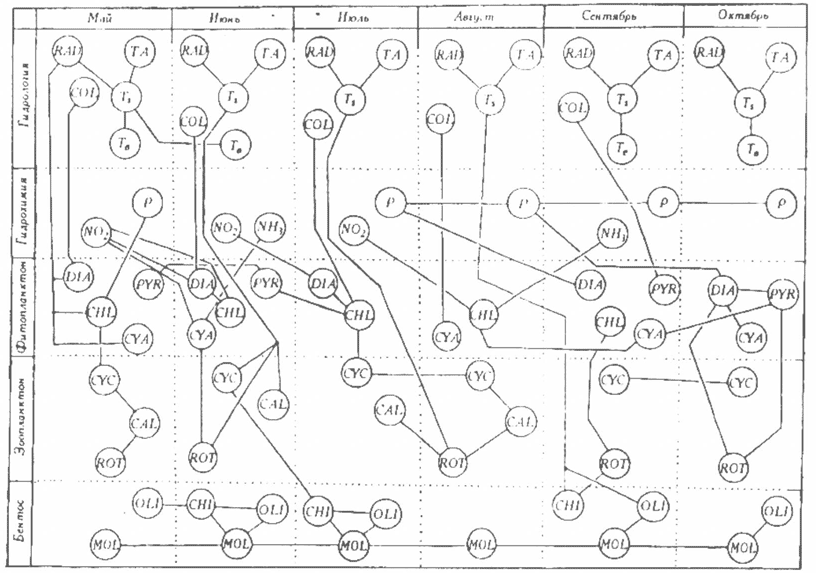
4. Связи в экосистеме на ст. 34 Куйбышевского водохранилища. Показаны только те связи, для которых коэффициент корреляции положителен и статистически значим при Р = 0,95.
Начнем рассмотрение с гидрологической части экосистемы. То, что температура поверхности воды (Ts) во всех случаях коррелирует с интенсивностью солнечной радиации (RAD) и температурой воздуха (ТА), достаточно тривиально. Но тот факт, что придонная температура воды в июне (Тв) зависит от поверхностной температуры (Ts) в мае, уже характеризует процесс весеннего прогрева водохранилища. Поверхностная и глубинная температура воды обнаруживают корреляционную связь только в мае, сентябре и октябре. Это позволяет отнести Куйбышевское водохранилище к стратифицированным водоемам с ярко выраженными периодами весеннего и осеннего перемешивания водных масс.
Для упрощения рис.4 из рассмотрения опущен режим растворенного кислорода и углекислоты и оставлены только корреляционные связи, относящиеся к биогенным элементам — азоту и фосфору. Характерна взаимная корреляция содержания минерального фосфора (Р) в воде с июля по октябрь, которая косвенно может свидетельствовать о том. что “весенний" фосфор в основной части уходит в донные отложения и транзитом в нижерасположенные водохранилища. “Остаточное” его количество в летне-осенний период, когда водообмен снижается, непрерывно участвует в биологическом круговороте.
Концентрация минерального фосфора в мае коррелирует только с численностью зеленых водорослей (CHL) в том же месяце, его содержание в июле и августе — с численностью диатомовых водорослей (DIA) соответственно в августе и октябре. Интересно, что из форм азота более многочисленными корреляции у фитопланктона оказались с содержанием в воде нитритов (NO2-), причем с их количеством в первой половине периода открытой воды. Из других форм азота содержание NH4+ в июне коррелировало с численностью сине-зеленых водорослей в том же месяце. Полученные корреляции уровня развития тех или иных групп водорослей с содержанием биогенных элементов — это первая информация такого рода относительно фитопланктона волжских водохранилищ. Насколько они устойчивы, можно определить, продолжив исследования массивов данных по другим станциям водохранилища. Несколько странной представляется корреляция численности зеленых водорослей (CHL) в августе с концентрацией аммонийного иона (NH4+) в сентябре.
Тот факт, что численность диатомовых (DIA), зеленых (CHL) и сине-зеленых (CYA) водорослей коррелирует с солнечной радиацией (RAD) только в мае, а во все остальные месяцы подобная связь отсутствует, свидетельствует о том, что лимитирующим фактором в развитии фитопланктона свет является только в весенний период. Возможно, что эта связь имеет опосредованный характер, когда изменения потока солнечной радиации определяют даты вскрытия водоема, интенсивность прогрева водных масс, условия стратификации - факторы, весьма значимые в развитии весенних альгоценозов. Обнаруживается связь развития фитопланктона с цветностью воды (COL). В мае и июне этот показатель коррелирует с численностью диатомовых (DIA), в июле — зеленых (CHL), в августе — сине-зеленых (CYA), a в сентябре - с численностью пирофитовых водорослей (PYR).
Интересно, что по связям внутри сообщества фитопланктона сезон активной вегетации водорослей четко делится на два периода: с мая до июля (в весенне-раннелетнем фитопланктоне) и с августа до октября (в летне-осеннем). Но эти связи немногочисленны.
В зоопланктоне зависимость биомассы основных групп от температуры воды проявляется только в июне. Позднее обнаруживается лишь корреляция температуры воды (Ts) в июле с биомассой коловраток (ROT) в августе. Внутри сообщества зоопланктона связи еще более слабые, чем в сообществе фитопланктона.
Между элементами зоопланктонного и фитопланктонного сообществ отмечаются связи в основном с отрицательными коэффициентами корреляции и находящиеся на границе статистической значимости (на рис. 4 они не изображены). Значима связь биомассы циклопоид (CYC) с численностью зеленых (CHL) водорослей в мае и июле, сине-зеленых (CYA) в сентябре. В октябре биомасса коловраток (ROT) связана с численностью и диатомовых (DIA), и пирофитоных (ROT). и зеленых (CHL) водорослей. Эти связи, очевидно, опосредованы, так как интенсивность развития той и другой группы организмов определяется скоростью весеннего прогрева и осеннего охлаждения водных масс.
Слабые связи, также не отмеченные на риссунке, оказались у бактериопланктона с другими компонентами и характеристиками экосистемы водохранилища.
Все это отражает, очевидно, относительно слабую структурированность планктона евтрофного водохранилища, что в известной степени согласуется с мнением о пелагических биосистемах как о системах со слабой внутренней организацией [15].
Между зоопланктоном и бентосом значимых корреляционных связей на ст. 34 Куйбышевского водохранилища также почти не обнаружено, Зато внутри бентосного сообщества эти связи очень сильны. Наиболее консервативным элементом всей экосистемы оказалась биомасса моллюсков (MOL), которая коррелирует не только сама с собой на протяжении всех шести месяцев наблюдения, но и с биомассой олигохет (OLI) в июне, июле, сентябре, октябре и хирономид (CHI) — в июне, июле.
Представление связей в водной экосистеме в форме, изображенной на рис. 4, применено не впервые. Аналогичный граф составлен для экосистемы оз. Дальнего по данным сорокалетних наблюдений — с 1935 по 1974 г. [3; с. 52, рис. 1.82], где число исследуемых характеристик было меньше, чем в Куйбышевском водохранилище, зато продолжительность непрерывных наблюдений была больше и велись они круглый год. При всей непохожести таких водоемов, как оз. Дальнее и Куйбышевское водохранилище, графы корреляционных связей дли них, построенные по одной и той же методике, имеют некоторое сходство как в области формирования температуры воды, так и в области взаимодействия фитопланктона с концентрацией минерального фосфора и азота.
Граф связей в экосистеме (см. рис. 4) представляет собой по существу развернутую во времени блок-схему имитационной модели водной экологической системы, построенной исключительно на эмпирических данных и не учитывающей законы сохранения вещества и энергии |5].
Заключение
Эксплуатация банка данных по Куйбышевскому водохранилищу еще только началась, и полученные результаты свидетельствуют скорее о доброкачественности исходного материала, чем о его полном использовании. Тот факт, что при достаточно разностороннем использовании данных не было получено абсурдных и явно противоречащих установившимся положениям современной гидробиологии результатов, свидетельствует о том, что созданный банк данных может служить основой для дальнейших изысканий. Часть выявленных связей имеет причинно-следственный характер, другие — статистический, а третьи заставляют задуматься и, по крайней мере, учитывать их при последующих исследованиях. В целом же в области изучения данных по Куйбышевскому водохранилищу сделан не больше чем первый шаг в стиле разведочного анализа [7]. В перспективе на основе собранных данных возможно построение экспертной системы [13] по определению характеристик водохранилища при различных внешних воздействиях, что может оказаться более эффективным, чем разработка классической имитационной модели водной экосистемы [14].
Литература
Глубокоуважаемые читатели! Оттиск статьи был извлечен из макулатурной древности и лист со списком литературы был утерян. Журнал из библиотеки ИЭВБ также был злостно похищен. Приносим свои извинения.
Однако, если кто-то не сочтет за труд выслать администраторам сайта нужную информацию, этот пробел будет немедленно восполнен.