© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic"
На главную страницу сайта
Сайт наш чаще посещай - будет выше урожай (Лозунг времен Н.С.Хрущева)
|
© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic" |
|
Опубликовано в сборнике "Количественные методы экологии и гидробиологии (памяти А.И. Баканова)", 2005.
![]() Скачать полный текст сборника в формате PDF (5.4 Мбайт)
Скачать полный текст сборника в формате PDF (5.4 Мбайт)
УДК (574.5 + 597):57.087.1
ОСНОВНЫЕ ИСТОЧНИКИ ОШИБОК В ГИДРОБИОЛОГИЧЕСКИХ И ИХТИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЯХ
А.И. Баканов, М.М. Сметанин, Н.М. Шихова
Институт биологии внутренних вод им. И. Д. Папанина РАН, 152742, пос. Борок, Ярославская обл., Некоузский р-н
Кратко изложены основные положения теории ошибок. Показаны основные источники ошибок, возникающих при количественной оценке пресноводного зообентоса, при анализе роста и возраста рыб.
Введение
“Errare humanum est” (Человеку свойственно ошибаться -лат.)
Обязательным компонентом разработки научных методов и конкретных методик должен быть раздел, показывающий, какие ошибки могут возникнуть при применении того или иного метода, как оценить их величину, какие существуют пути устранения, уменьшения или компенсации этих ошибок [5, 6, 26, 31]. К сожалению, в гидробиологии и ихтиологии теория ошибок разработана недостаточно, многие исследователи обходят этот вопрос стороной, зачастую не проводится даже элементарная статистическая обработка исходного материала. Выводы авторов многих публикаций при внимательном рассмотрении оказываются неубедительными, а практические рекомендации – иногда ошибочными.
Ранее [32] авторы достаточно подробно рассмотрели основные положения теории ошибок, поэтому здесь они будут изложены очень кратко. Цель данной работы - выявление основных источников ошибок, возникающих в некоторых гидробиологических и ихтиологических исследованиях, а также иллюстрация их соответствующими примерами. Вопросы оценки ошибок и возможности их уменьшения – тема следующей публикации.
Изучаемые исследователем объекты и их отдельные свойства подлежат измерению. Измерение – совокупность действий, выполняемых с помощью средств измерений и имеющих целью нахождение числового значения измеряемой величины, выраженного в принятых единицах измерения. Для количественной характеристики какого-либо явления, объекта или процесса надо построить понятие особого рода – показатель. Он должен давать количественную характеристику явления в единстве с его качественной определенностью. Показатель состоит из модели качественной стороны явления, устанавливающей, что, где, когда и каким образом подлежит измерению (качественный реквизит), и численного выражения явления в конкретных условиях места и времени (количественный реквизит) [34]. Как правило, каждому гидробиологическому и ихтиологическому понятию должен соответствовать определенный показатель, ибо вводить понятия, которые нельзя охарактеризовать количественно, нецелесообразно. Известно, что всякое явление или процесс может иметь много различных сторон или аспектов. Лишь сравнительно простое явление можно охарактеризовать одним показателем. Сложные явления характеризуются целой системой показателей, каждый из которых измеряет какой-либо один аспект явления. На основании их затем возможно построение комплексных показателей.
Современный количественный подход к решению большинства гидробиологических задач предполагает не только измерение изучаемых параметров, но и оценку точности исследований. Анализ проводится, как правило, выборочным методом. Из изучаемой совокупности объектов (обычно ее называют генеральной совокупностью) отбирают определенным образом некоторую часть, и на основании этой части (выборки) делается заключение о генеральной совокупности. Это заключение производится с определенной вероятностью и содержит некоторую ошибку, которая может иметь несколько составляющих.
Качество любого измерения характеризуется его погрешностью (или точностью). Погрешностью измерения называется отклонение его результата от истинного значения измеряемой величины. Под точностью понимают качество измерения, отражающее близость его результата к истинному значению измеряемой величины. Точность и погрешность тесно связаны между собой. Чем больше точность, тем меньше погрешность, т. е. точность оценивается через погрешность. На практике в биологической литературе вместо термина "погрешность" чаще используют слово "ошибка".
По форме погрешности разделяются на абсолютные и относительные. В зависимости от характера отклонений ошибок от истинного значения погрешности делятся на систематические, случайные и промахи. В зависимости от источника возникновения можно выделить ошибки объекта, приборные или инструментальные, методические, от внешних источников и личные. Среди разнообразных методических ошибок следует обратить особое внимание на ошибки концептуальные, которые во многих случаях могут быть самыми опасными. Концепция (от латинского conceptio – понимание) – определенный способ понимания, трактовки какого-либо объекта, явления, процесса, руководящая идея для их изучения. Сюда относятся, например, неправильная трактовка объема понятия – его слишком расширительное или суженное толкование, применение неадекватных методов статистической обработки. Попытки дать "точную" количественную оценку понятиям, логически строго не определенным, расплывчатым или интуитивным создают только видимость математического обоснования выводов.
К такого рода ошибкам может привести некритическое применение выводов, полученных в лабораторных экспериментах, к явлениям и процессам в естественных экосистемах. В функционировании водных экосистем огромную роль играет пространственный и временной масштаб, который не может быть соблюден в лабораторных условиях. В гидробиологических и ихтиологических исследованиях встречаются практически все виды ошибок.
Примеры из области изучения пресноводного зообентоса. Зообентос - экологическая группа, объединяющая животных, основную часть жизни обитающих на поверхности или в толще грунта. Разными исследователями объем этого понятия определяется неодинаково, что приводит к концептуальным ошибкам. На мелководных участках водоемов на грунте растут высшие водные растения, на которых, между которыми и внутри которых обитают животные. Одни гидробиологи всех этих животных относят к бентосу, другие выделяют их в особую экологическую группу, называемую зоофитосом, фитофильной фауной или зооперифитоном. Нечеткость определения термина "бентос" затрудняет количественную оценку его параметров. Поэтому списки видов и прочие характеристики бентоса одного водоема, представленные различными исследователями, придерживающимися разного подхода к этому вопросу, могут сильно различаться.
Первая задача, возникающая при изучении бентоса нового водоема, - установление его видового состава, что предусматривает проведение количественных и качественных сборов. Зообентос - удивительно разнообразная в таксономическом отношении экологическая группа, включающая 17 классов животных (без учета простейших и паразитических форм). При этом, например, класс насекомых в бентосе представлен 11 отрядами, а отряд Diptera - 22 семействами, тогда как зоопланктон, представлен в основном ракообразными и коловратками.
Отсюда вытекает очень низкая степень изученности зообентоса в рассматриваемом плане. Например, в монографии [28] список видов включает представителей всего семи классов бентосных животных, причем из 11 отрядов насекомых приведено всего два, а из 22 семейств отряда двукрылых всего одно - хирономиды. По многим группам бентосных беспозвоночных нет определителей и, очевидно, отсутствуют специалисты. Списки видов и показатели биоразнообразия многих водоемов в значительной степени отражают не столько реальное богатство фауны, сколько степень изученности водоема.
Положение осложняется тем, что в ходе практической работы с собранным материалом значительная часть пойманных особей не может быть определена до вида. В первую очередь это касается личиночных стадий водных насекомых, которые определяются до личиночной формы, группы видов, группы родов и т. д. , однако и у других водных организмов, например, у олигохет, составляющих часто более 50% бентоса, основная часть особей представлена неполовозрелыми экземплярами, тоже в большинстве случаев не определяемых до вида [4].
При сравнении фаун разных районов или населения разных биотопов исследователь, как правило, не знает полного числа видов изучаемых животных (S), их составляющих. Имея дело с выборками животных, он может определить только число видов, которые попали в эти выборки. При увеличении объема выборки точность оценки S увеличивается. Прямое сравнение числа видов в выборках, зависящее от объема последних, может искажать истинные различия в богатстве видами сравниваемых фаун и населения. Для более точного сравнения различных водоемов (участков, биотопов...) по числу видов необходимо выяснить, как изменяется величина S при изменении объема собранного материала (количества и размера проб, числа собранных особей). Эта зависимость аппроксимируется различными теоретическими распределениями [24], сравнение параметров которых позволяет делать более обоснованные выводы о видовом богатстве.
Зообентос служит кормовой базой рыб-бентофагов, составляющих основу уловов в пресноводных водоемах. В этом случае особенно важно получать точные данные о состоянии зообентоса, используемые в дальнейших трофологических расчетах. Можно выделить пять основных источников ошибок в оценке кормовой базы рыб [2, 3].
1. Использование для сбора бентоса неподходящих по конструкции или неудовлетворительно работающих дночерпателей. Судя по публикациям, примерно 75% отечественных исследователей для сбора материала применяют дночерпатель Петерсена, остальные - различные модификации дночерпателя Экмана, а также несерийные, специально изготовленные приспособления. Известно, что эффективность различных устройств на разных типах грунтов и по отношению к разным группам донных организмов неодинакова, стандартные дночерпатели могут в несколько раз занижать оценку зообентоса. Дночерпатели плохо или совсем не учитывают организмы нектобентоса, куда относятся в первую очередь мизиды, гаммариды и другие ракообразные, играющие существенную роль в кормовой базе, особенно южных водоемов. Чрезвычайно низка точность учета в водоемах крупных моллюсков - дрейссены и унионид, вследствие чего создается впечатление об огромных колебаниях их биомассы. Например, в Цимлянском водохранилище на песчаном грунте среднелетняя биомасса бентоса, в которой доминировали моллюски, в 1960-1971 гг. изменялась от 1.2 до 1035 г/м2 [23]. Для сравнения уровня развития бентоса в разных водоемах используются показатели так называемого “дночерпательного бентоса”, который составляет только часть общего бентоса.
2. Ошибки как результат фиксации и методики разборки материала. При бентосных съемках водоема количество собранного материала бывает так велико, что для дальнейших исследований его приходится фиксировать в растворе формалина или спирта. При фиксации масса организмов меняется, причем величина этих изменений зависит от многих факторов, у некоторых организмов она может превышать 50%. Неучет этого фактора может привести к значительным систематическим ошибкам в оценке биомассы бентоса. Путем изучения соотношения массы живых и фиксированных организмов в конкретных условиях определенного водоема необходимо найти поправки к величинам фиксированной биомассы, однако, обычно это не делается.
Один из самых трудоемких этапов бентосных работ - выделение организмов из грунта. Эта операция выполняется вручную после промывки проб через сито из мельничного газа с ячеей разного размера для отделения частиц грунта. В зависимости от размеров животных и диаметра ячеи часть организмов теряется. Поскольку потери идут за счет более мелких организмов, это приводит не только к занижению показателей обилия, но и к искажению значений средней массы особи, показателей видового разнообразия и т. п.
Выбор размера ячеи зависит от того, какую фракцию зообентоса нужно уловить, причем выделяют от 2 до 5 размерных фракций. Наиболее часто изучается макробентос, а именно, организмы, которые можно достаточно полно выбрать из грунта без применения оптики, они имеют размер от 2-3 мм и более и задерживаются газом № 9-11. В некоторых водоемах макробентос составляет менее половины общего бентоса, остальная часть которого теряется при промывке [2].
3. Ошибки от неправильно планирования сборов. Обычно пробы бентоса берутся на гидробиологических разрезах или в местах, которые, по мнению исследователя, достаточно типичны для водоема. По собранному таким образом материалу невозможно достоверно судить о величине генеральных параметров, которые должны оцениваться по выборке. Теория выборочного исследования редко используется при бентосных исследованиях, что приводит к смещенным оценкам генеральных параметров.
4. Статистические ошибки. При гидробиологических исследованиях единичные данные, характеризующие состояние бентоса в отдельных точках водоема в момент исследования, не представляют интереса. Ценность имеют только средние данные, характеризующие бентос определенного участка или всего водоема за определенный срок. В силу естественной вариабельности плотности бентоса в разных точках водоема средние данные всегда будут иметь некоторую статистическую ошибку, называемую также ошибкой репрезентативности. Ее необходимо оценивать, что очень часто не делается. Без знания величины этой ошибки нельзя рассчитать доверительный интервал средних значений, невозможно статистическое сравнение данных разных исследователей. Величина ошибки зависит от степени вариабельности бентоса и количества собранных проб, а также от объема материала, входящего в пробу [38]. В силу трудоемкости бентосных работ на водохранилище отбирается сравнительно небольшое количество проб. Например, при изучении бентоса Рыбинского водохранилища разные исследователи отбиралось от 40 до 175 проб, в последнем случае стандартная ошибка составила ~ 20% при площади одной пробы 250 см2. Такая величина ошибки для полевых работ считается удовлетворительной, но обычно она превышает 50%.
Поскольку плотность различных видов сильно варьирует, то для того, чтобы в одном и том же месте получить среднюю биомассу с ошибкой < 30%, для одного вида достаточно взять, например, шесть проб, а для другого – 144 [43].
5. Ошибки в оценке доступности организмов для рыб. Доступные рыбам кормовые организмы составляют только часть общего бентоса, неодинаковую в разных водоемах. Доступность кормовых организмов зависит от их размеров, характера распределения, видового и возрастного состава рыб, а также ряда других факторов. Известно, что крупные моллюски, дающие основную часть биомассы бентоса во многих водоемах, потребляются рыбой значительно слабее, чем хирономиды и олигохеты. По мнению А.Ф. Алимова [1] рыбы могут использовать ~ 30% продукции крупных моллюсков.
Организмы мейобентоса входят в рацион главным образом молоди рыб. Их роль в питании взрослых рыб значительно меньше, чем в продукции бентоса, поскольку это высокодисперсный и, следовательно, энергетически менее выгодный корм.
Особенности вертикального распределения организмов в толще грунта существенно влияют на их доступность. Биомасса бентоса, расположенная глубже 10 см на илистых грунтах считается временно недоступной [9]. В Рыбинском водохранилище на илистых грунтах за счет этого ~ 23% биомассы бентоса недоступно для рыб, а в Водлозерском водохранилище > 90% мотыля находится в глубоких слоях грунта [15]. На песчаных грунтах большая часть бентоса может быть недоступна для рыб.
Как показали эксперименты В.С. Ивлева [17], на обеспеченность рыб пищей сильное влияние оказывает характер ее распределения на дне. Чем более агрегированы организмы бентоса, тем более доступны они для рыб, увеличение агрегированности эквивалентно увеличению биомассы корма [17]. Агрегированность бентоса различных водоемов сильно различается, наблюдаются и ее сезонные колебания.
Важнейшая характеристикой организмов любого систематического и трофического уровня - величина производимой ими продукции. Она лежит в основе энергетических и трофологических расчетов, необходима для правильной оценки эффективности функционирования всех звеньев экосистемы и имеет важнейшее прикладное значение. Точность расчета продукции донных организмов остается, как правило, неизвестной; априори можно утверждать, что она весьма низкая. Например, сравнивая в экспериментальных условиях "фактическую" продукцию личинок хирономид Chironomus dorsalis с теоретически ожидаемой ее величиной, полученной методом суммирования суточных приростов, в первом варианте опытов получили, что расчетная продукция составляла 88.1% фактической, во втором – 130.4% [19].
В.И. Дулепов [16], изучая в экспериментальный условиях продукцию озерного бокоплава Anisogammarus kygi, оценил относительную ошибку определения продукции в 20.3%, а P/B коэффициента – 42.1%. В полевых же условиях, когда на ошибки теоретических продукционных моделей накладываются огромные ошибки репрезентативности оценки численности и биомассы многовидовых сообществ животных, точность продукционных расчетов будет значительно ниже. Например, погрешность определения такого важного продукционного показателя, как P/B коэффициент, в полевых условиях может быть > 100% [30]. Канадские исследователи [41] моделировали расчет продукции одного вида бентосных организмов разными методами и нашли, что систематические ошибки составляют 50-60%, а случайные – 30-500%.
Накопление ошибок при рыбопродукционных расчетах может привести к совершенно противоположным выводам. Например, рассчитывая дополнительную рыбопродукцию, которая может быть получена в волжских водохранилищах за счет использования резервов зоопланктона М.Л. Пидгайко [25] получила для Рыбинского водохранилища отрицательную величину, т. е. зоопланктон этого водоема используется полностью. По данным же сотрудников ИБВВ РАН [8] в Рыбинском водохранилище рыба выедает < 3% продукции зоопланктона, т. е. имеются значительные резервы роста рыбопродукции за счет более полного его использования.
Примеры из области изучения роста и возраста рыб.
Использование неоптимальных показателей роста рыб (ПРР). При анализе роста нужно отличать данные, получаемые при исследовании совокупности особей известного возраста, данные одномоментной выборки и информацию о росте отдельных особей [22]. Оперируя с изменениями роста совокупности особей известного возраста, необходимо учитывать репрезентативность выборки, а при исследовании одномоментного облова, кроме того - стабильность роста различных поколений.
Несомненно, что изучение семейства кривых, описывающих рост отдельных особей, дает наиболее полную исходную информацию, которую в частности, можно использовать и для оценки точности показателей роста. Например, для восьми меченых особей леща с заведомо известным возрастом 6+, выращиваемых в пруду с октября 1972 г. по октябрь 1973 г. среднее значение удельной скорости роста составило 0.12 длины тела в год, а возможная погрешность с вероятностью 0.95 равна 0.01 длины тела.
Часто имеется информация о росте рыб, получаемая в результате анализа выборок, производимых в последовательные моменты времени, т.е. когда прослеживается рост не отдельных особей, а определенного годового класса. В таком случае нет возможности определения интервальных показателей роста у отдельных особей, но статистические погрешности этих показателей можно рассматривать как ошибки косвенных измерений.
При сопоставлении скорости роста, выявлении его периодичности, при изучении экологии отдельных видов рыб используются различные ПРР. По признаку времени их можно разделить на две группы - моментные и интервальные. К моментным относятся показатели, дающие количественное описание роста на определенный момент времени, например, размер (длина, масса тела и т.д.) в данный момент времени. К интервальным относятся показатели, дающие количественную характеристику явления за определенный период времени. ПРР могут быть также разделены на абсолютные и относительные. Абсолютные (абсолютный прирост, размер тела в данный момент времени, средняя абсолютная скорость роста, абсолютное ускорение и др.) характеризуют рост всего тела. Относительные (относительная и удельная скорость роста, константа роста, коэффициент роста, относительное ускорение роста и др.) относятся к показателям, характеризующим рост единицы длины или массы тела.
1. Использование “характеристики” вместо абсолютной скорости роста рыб. Для описания неравномерности роста В.В. Васнецовым [11] была предложена “характеристика роста” H, размерность которой совпадает с размерностью абсолютной скорости роста V . Например, если длина тела Li измеряется в миллиметрах, а возраст рыбы в годах, то размерность и скорости, и “характеристики роста” - мм/год, оба эти показателя можно считать мерой экстенсивности роста.
Показано [29], что систематическая ошибка “характеристики роста” сравнительно мала только в небольшой окрестности точки Li+1/Li = 1 и возрастает с увеличением этого отношения. Причем, наиболее быстрое увеличение функции (на 30.7%) наблюдается в часто встречающейся на практике области значений Li+1/Li - от 1 до 2. Поэтому при использовании H вместо V, помимо статистической ошибки, как правило, появляется и значительная систематическая погрешность.
Например, у выращенной в прудах густеры средняя длина тела в возрасте 1 год составляла 47 мм, в возрасте 2 года - 92 мм, а погрешность “характеристики роста” - > 30%. Еще большие систематические ошибки получаются при увеличении отношения Li+1/Li. Так, расчеты показывают, что при увеличении средней длины молоди щуки в течение 132 сут. с 11.9 до 138.7 мм [27] систематическая ошибка “характеристики роста” будет > 75 %.
2. Использование неоптимальных показателей относительной скорости роста. Для изучения интенсивности роста используются относительные показатели. Их критический анализ [14, 22] позволил установить, что наиболее точная оценка относительного прироста в единицу времени - удельная скорость роста [33]. Интересно сопоставить с ней часто используемые показатели относительной скорости роста по формулам Майнота [40] и Броди [36]. Показано, что различие между величинами этих показателей зависит от отношения Li+1/Li [29].
Для рыб наибольшая величина этого отношения при одинаковых интервалах дискретности наблюдается в первое время после выклева. При этом даже за время 20-30 суток длина рыбы может увеличиться в несколько раз. Так, для леща нерестово-выростного хозяйства в р. Бирючек средняя длина тела на этапе развития А составляла 4.8 мм, а через месяц на этапе развития G - 25 мм [12], в этом случае систематическая погрешность относительной скорости роста при использовании индекса Майнота - > 145 %, а при применении индекса Броди - ~ 20 %.
Еще большие ошибки получаются при увеличении отношения Li+1/Li . Так, при выращивании годовиков карпа в прудах стационара полевых и экспериментальных работ ИБВВ РАН "Сунога" в течение 130 сут. их средняя масса увеличилась от 11 до 338 г. Систематическая погрешность индекса Броди составила более 65% , индекса Майнота - более 660%. Следовательно, для определения средней относительной скорости роста можно пользоваться формулами Майнота и Броди [36] только, если отношение Li+1/Li несущественно > 1.
Показатели относительной скорости роста в форме, предложенной Л.С. Бердичевским [7] и В.Л. Брюзгиным [10], - функции, пропорциональные абсолютной скорости роста с коэффициентами пропорциональности соответственно 1/Ln и 1/L0 . Здесь Ln - длина тела рыбы предельного возраста, а L0 - на первом году жизни. Эти показателя представляют собой масштабноизменённую абсолютную скорость роста. Естественно, что они в большинстве случаев отличаются от удельной скорости роста не только по величине, но и по характеру изменения.
Интенсивности линейного роста популяции по Балону [35] выражается усреднённым коэффициентом относительного прироста в форме Майнота [40], т.е. при интенсивном росте имеет большую систематическую погрешность, достигающую десятков и сотен процентов. Для количественного анализа быстроты изменения удельной скорости роста целесообразно применять относительное ускорение роста [29]. Если оно положительно, то свидетельствует об ускорении роста единицы длины тела рыбы, если отрицательно, о замедлении относительного роста.
Использование предельных погрешностей вместо среднеквадратических. Предельную абсолютную косвенную ошибку функции можно найти по формуле:
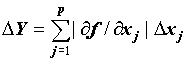
 (1)
(1)
При этом предельная косвенная ошибка понимается в вероятностном смысле, так как ошибки аргументов оцениваются для избранного уровня вероятности. Кроме того, предполагается самая невыгодная ситуация, когда все частные погрешности имеют одинаковые знаки.
Очевидно, что при xj ¹ xj+1, xj > 0 рассматриваемые функции непрерывны относительно всех аргументов и имеют производные. Применяя к ним соотношение (1) можно получить формулы предельных среднеквадратических ошибок показателей роста с учетом погрешности интервала дискретности. Однако применение этих формул приводит, вообще говоря, к завышению результирующей ошибки. Кроме предельной ошибки, имеет смысл вычислить еще и среднеквадратическую погрешность, величина которой часто в 1.5-2 раза меньше [29] .
Неучет корреляционной связи между аргументами ПРР. Для линейной функции :
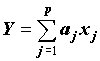 (2) ,
(2) ,
стандартная ошибка функции D Y выражается через стандартные ошибки измерений D xj и коэффициенты корреляции между ними rjk следующим равенством [13, 18]:
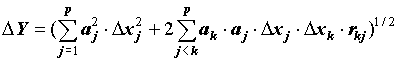 (3) .
(3) .
В тех случаях, когда ошибки измерений независимы или некоррелированы , rjk = 0 и формула (3) упрощается:
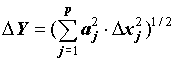 (4) .
(4) .
Из взятых показателей линейная функция своих аргументов - абсолютный прирост Li+1 - Li. При независимости ошибок D Li и D Li+1:
D
Y = ((D
Li+1)2+(D
Lj)2)1/2 (5) .
Если функция Y = f(x1, x2,...xj,..,.xp) линеаризуема с достаточной степенью в рассматриваемой области изменения аргументов, то ее стандартная ошибка D Y выражается через стандартные ошибки измерений и коэффициенты корреляций между ними rjk следующим приближенным равенством:
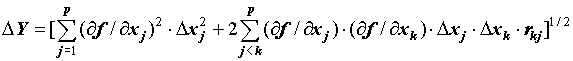
![]() (6)
(6)
Формула (6) была применена к наиболее распространенным в экологии показателям: росту, упитанности, пространственному распределению, плодовитости, убыли, смертности. При анализе полученных зависимостей, в частности для показателя относительной скорости роста рыб, было выявлено, что в формулу погрешности слагаемые входят с разными знаками, что приводит к уточнению величины среднеквадратической погрешности (СП). Например, для показателя коэффициента упитанности рыб по Кларк [22] показано, что слагаемые, содержащие в качестве множителя коэффициент корреляции между переменными могут вносить существенный (до 30 %) вклад в результирующую СП.
Для ряда выведенных формул (относительной и абсолютной скоростей роста, коэффициентов упитанности по Кларк и Фультону, коэффициентов плодовитости, вариации, агрегированности авторами были написаны программы на языке PASCAL и выполнены вычисления на ЭВМ. В результате их применения установлено, что для некоторых показателей зависимость СП от коэффициента корреляции r между аргументами весьма существенная. Например, при изменении r от 0 до 1 между средней длиной тела в последовательные моменты времени от 0 до 1 величина СП удельной скорости роста возрастает более чем в 10 раз. Напротив, при изменении r между средним и дисперсией от -1 до 1 величина СП коэффициента вариации уменьшается почти в 6 раз.
Недоучет нелинейности поведения показателей. Показатели, используемые в ихтиологических исследованиях, можно интерпретировать как функции нескольких переменных. В ряде случаев в рассматриваемом диапазоне изменения аргументов эти функции достаточно хорошо линеаризуемы, а составляющая среднеквадратической погрешности, обусловленная нелинейностью функции, относительно мала. Так, вычисления показывают, что для средней удельной скорости роста нескольких десятков одновозрастных особей окуня, выращиваемых в прудах стационара полевых и экспериментальных работ ИБВВ РАН “Сунога”, при изменении средней длины тела от 50± 1 мм (возраст 1+) до 120± 1мм (возраст 2+) указанная составляющая не превышает нескольких процентов.
Если же выбранный индекс в наблюдаемой области изменения переменных существенно нелинеен, то поправка, обусловленная этим, может быть достаточно велика. Например, для наиболее широко применяемого показателя агрегированности Сведберга [42], иногда называемого коэффициентом или показателем дисперсии, D = σ2/m, где σ2 – дисперсия, m - среднее арифметическое значение, поправка на нелинейность может достигать несколько сотен процентов от величины линейной составляющей погрешности.
Ошибки определения возраста рыб. Общая погрешность определения показателей роста в значительной степени зависит от точности определения возраста. В работе М.В. Мины [21] обоснована необходимость и намечены пути оценки точности используемых методик и, в первую очередь, наиболее простого и распространённого способа - визуального определения возраста по регистрирующим структурам.
1. Систематическая погрешность. При определении возраста рыб систематические ошибки увеличивают или уменьшают истинное число прожитых лет. Значительные систематические погрешности определения возраста рыб (год или больше) могут возникать из-за особенностей регистрирующих структур и в результате личных систематических погрешностей операторов.
Первое годовое кольцо на чешуе ельца из водоемов Англии закладывается, например, в конце второго или начале третьего года жизни [39]. М.В. Мина [21] также отмечал образование на чешуе у большинства исследованных им особей микижи р. Камчатки первой зоны сближенных склеритов лишь в конце второго или в начале третьего года жизни. Показано также несоответствие количества годовых зон на чешуе заранее известному возрасту всех исследованных особей угря [37]. Следовательно, число годовых колец на чешуе может не соответствовать истинному возрасту рыб.
Личные систематические ошибки оператора могут заключаться как в преимущественном принятии дополнительных колец за годовые, так и годовых за дополнительные. При этом наблюдается смещение центра распределения возрастных оценок, даваемых оператором, от истинного возраста. Интересно, что систематическая ошибка может появляться, начиная с определённого возраста рыб. Так, при анализе восемью операторами чешуи 15 особей плотвы с заведомо известными возрастами 3+, 4+, 5+, выращенной в прудах стационара полевых и экспериментальных работ ИБВВ РАН “Сунога” (операторам сведения о возрасте особей не сообщались), модальные значения распределения возрастных оценок совпали с истинным возрастом, а для особей возраста 5+ мода и истинное значение возраста смещены на 1 год. При этом в 75% определений наблюдалось систематическое занижение возраста на 1-2 года. По видимому, это связано с тем, что пятое годовое кольцо менее резко отличается от дополнительных, чем предыдущие годовые кольца.
Помимо анализа чешуи рыб с заведомо известным возрастом, выявление и устранение систематических ошибок связано с использованием и других регистрирующих структур (отолитов, костей) , а также иных, более совершенных методик.
2. Случайная погрешность. Случайная (статистическая) ошибка при визуальном определении возраста рыб может быть вызвана различными причинами - степенью чистоты чешуи, освещенностью, состоянием оператора и т.д. Как указали Е.А Криксунов и А.Е. Бобырев [20], наличие случайных ошибок в определении возраста рыб может приводить к возникновению систематических погрешностей в оценке возрастной структуры уловов и, как следствие, к погрешности оценки запаса.
О величине случайной погрешности можно судить, определяя возможный разброс или воспроизводимость оценок возраста одних и тех же особей разными операторами. При этом чем меньше разброс, тем, в среднем, меньше случайная ошибка. Так, при визуальном подсчете годовых колец чешуи плотвы выращенной в прудах с заведомо известным возрастом 3+, 4+, 5+, а также леща и густеры Рыбинского водохранилища процент воспроизводимости показаний разных операторов варьирует в широких пределах: для плотвы - 13-51%, для леща - 0-50%, для густеры - 30-55% соответственно. Гораздо больше он оказался при неоднократном определении возраста густеры Рыбинского водохранилища одним и тем же опытным оператором (30-95%). Следовательно, у данной выборки одни и те же образования чешуи густеры разные операторы различно интерпретируют чаще, чем один оператор, неоднократно просматривая их даже через длительные промежутки времени.
Выводы. При планировании каждого исследования необходимо заранее предусмотреть возможные источники разных видов ошибок и их возможные величины. Особое внимание следует уделить тем ошибкам, которые в дальнейшем не могут быть исправлены или уменьшены (ошибкам репрезентативности, концептуальным ошибкам и т.д.). Предварительно необходимо выбрать обоснованную систему измеряемых и рассчитываемых показателей, рассмотреть их возможное поведение в разных диапазонах величин и имеющихся экологических ситуациях, найти формулы для вычисления их ошибок, а также установить, какая изучаемая характеристика вносит наибольший вклад в общую ошибку результатов. В случае превышения ожидаемой ошибки допустимой для данного типа исследований необходимо рассмотреть возможности ее уменьшения. Выводы должны соответствовать точности исследований.
СПИСОК ЛИТЕРАТУРЫ
1. Алимов А.Ф. Соотношения между трофическими уровнями в сообществах пресноводных животных // Журн. общ. биол. 1983. Т. 44. № 4. С. 435-445.
2. Баканов А.И. О репрезентативности данных по кормовой базе рыб-бентофагов // Вопр. ихтиологии. 1979. Т. 19. Вып. 6(119). С. 1133-1136.
3. Баканов А.И. Зообентос водохранилищ и использование его рыбами // ИБВВ АН СССР. 1989. 37 с. Деп. в ВИНИТИ 12.05.89 г., № 3169-B89.
4. Баканов А.И. Использование характеристик разнообразия зообентоса для мониторинга состояния пресноводных экосистем // Мониторинг биоразнообразия. М.: ИПЭЭ РАН, 1997. С. 278-282.
5. Баранов Ф.И. К вопросу о биологических основаниях рыбного хозяйства // Избр. тр. М.: Пищ. Пром-сть, 1971. Т. 3. С. 12-56.
6. Баранов Ф.И. О точности вычислений // Избр. тр. М.: Пищ. Пром-сть, 1971. Т. 3. С. 205-213.
7. Бердичевский Л.С. Биологические основы рационального использования рыбных запасов: Автореф. дис. ... докт. биол. наук. М.: ВНИРО, 1964. 36 с.
8. Биологические процессы во внутренних водоемах. Борок: Ин-т биол. внутр. вод АН СССР, 1981. 44 с.
9. Боруцкий Е.В. Вертикальное распределение биомассы бентоса в толще иловых отложений в некоторых подмосковных озерах // Зоол. журн. 1940. Т. 19. Вып. 2. С. 240-256.
10. Брюзгин В.Л. Методы изучения роста рыб по чешуе, костям и отолитам. Киев: Наукова думка, 1969. 188 с.
11. Васнецов В.В. Опыт сравнительного анализа линейного роста сем. карповых // Зоол. журн. 1934. Т. 13. Вып. 3. С. 540-584.
12. Васнецов В.В., Еремеева Е.Ф., Ланге Н.О. и др. Этапы развития промысловых полупроходных рыб Волги и Дона // Тр. Ин-та морфол. животн. АН СССР. М.: Наука, 1957. Вып. 16. С. 7-76.
13. Вентцель Е.С. Теория вероятностей. М.: Наука, 1964. 76 с.
14. Винберг Г.Г. Скорость роста и интенсивность обмена у животных // Успехи соврем. биол. 1966. Т. 61, Вып. 2. С. 274-293.
15. Гордеева-Перцева Л.И. Бентос Водлозера и использование его рыбами // Вопросы гидробиологии водоемов Карелии. Петрозаводск: Карельское кн. изд-во, 1964. С. 131-148.
16. Дулепов В.И. Оценка точности расчета продукции на примере озерного бокоплава Anisogammarus kygi // Математические модели популяций. Владивосток: Ин-т автоматики и процессов управления, 1979. С. 50-53.
17. Ивлев В.С. Экспериментальная экология питания рыб Киев: Наукова думка, 1977. 272 с.
18. Кондрашихин В.Т. Теория ошибок и ее применение к задачам судовождения. М.: Транспорт, 1969. 256 с.
19. Константинов А.С., Нечваленко С.П. О точности определения продукции хирономид методом суммирования суточных приростов // Гидробиол. журн. 1968. Т. 4. № 6. С. 71-82.
20. Криксунов Е.А., Бобырев А.Е. Когортный анализ в свете имитационных экспериментов // Вопр. ихтиологии. 1992. Т. 32. Вып. 4. С. 55-62.
21. Мина М.В. Способ объективизации и уточнения оценок возраста рыб, в частности, микижи р. Камчатки // Вопр. ихтиологии.1973. Т.13. Вып.1(78). С.109-118.
22. Мина М.В., Клевезаль Г.А. Рост животных. М.: Наука, 1976. 291 с.
23. Мирошниченко М.П. Многолетняя динамика и продукция зообентоса Цимлянского водохранилища // Тр. Волгогр. отд. ГосНИОРХ. 1974. Т. 8. С. 304-325.
24. Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 288 с.
25. Пидгайко М.Л. Расчет дополнительной рыбопродукции Волжского каскада за счет резервов зоопланктона // Изв. ГосНИОРХ. 1978. Т. 139. С. 112-114.
26. Поддубный А.Г. Значение работ по оценке точности результатов биологических исследований // Оценка погрешностей методов гидробиологических и ихтиологических исследований. Рыбинск: Ин-т биол. внутр. вод АН СССР, 1982. С. 3-8.
27. Пупырникова А.В. Сезонные изменения в питании и росте молоди щуки // Тр. ВНИИ рыб. хоз-ва и океаногр. М., 1953. Т. 24. С. 338-345.
28. Рыбинское водохранилище и его жизнь. Л.: Наука, 1972. 364 с.
29. Сметанин М.М. Погрешности количественных показателей роста рыб // Оценка погрешностей методов гидробиологических и ихтиологических исследований. Рыбинск: Ин-т биол. внутр. вод АН СССР, 1982. С. 43-62.
30. Сметанин М.М., Баканов А.И. О статистической и систематической ошибках P/B коэффициента // Проблемы экологии Прибайкалья. Иркутск: 1979. Ч. 1. С. 167-168.
31. Терещенко В.Г., Решетников Ю.С. О значении работ по оценке погрешностей результатов в экологических исследованиях // Теоретические проблемы экологии и эволюции (Третьи Любищевские чтения). Тольятти: Ин-т экол. волж. бассейна РАН, 2000. С. 198-208.
32. Терещенко В.Г., Сметанин М.М., Баканов А.И. Методы оценки погрешностей в экологических исследованиях // Оценки погрешностей методов гидробиологических и ихтиологических исследований. Рыбинск: Ин-т биол. внутр. вод АН СССР, 1982. С. 9-23.
33. Шмальгаузен И.И. Определение основных понятий и методика исследования роста // Рост животных, М.; Л.: Биомедгиз, 1935. С. 6-8.
34. Эдельгауз Г.Е. Достоверность статистических показателей. М.: Статистика, 1977. 278 с.
35. Balon E.K. On relative indexes for comparison of the growth of fishes // Vestnik Cs. spol. Zool., 1964. Bd 28 (4). S.11-21.
36. Brody S. Bioenergetics and growth. N.Y.: Hafner, 1945. 1023 p.
37. Сiepielewski W. Okreslanis wieku wegwrra (Anguilla anguilla L.) // Rosz. nauk. rol. 1975.V. 21. № 2. P. 27-33.
38. Elliott J.M. Some methods for the statistical analysis of samples of the benthic invertebrates // Freshwater Biological Association. Sci. Publ. 1971. № 25. 144 p.
39. Mathews C.P., Williams W.P. Growth and annual check formation of dace, Leuciscus (L.) // J. Fish. Biol. 1964. V. 4. № 3. P. 363-367.
40. Minot C.S. The problem of age, growth and death. L.: Murray, 1908. 280 p.
41. Morin A., Mousseau T.A., Roff D.A. Accuracy and precision of secondary production estimates // Limnol., Oceanogr. 1987. V. 32. № 6. P. 1342-1352.
42. Svedberg T. Ett bidrag till de stastistika metodernas användning inom växtbiologien // Svensk. bot. Tidskrift. 1922. V. 16. № 1. P. 1-8.
43. Watanabe N.C., Harada S. Some problems in sampling stream fauna, with particular reference to the variability of samples // Jap. J. Limnol. 1976. V. 37. № 2. P. 47-58.
Main sources of errors meeting in hydrobiological and ichthyological researches
A.I. Bakanov, M.M. Smetanin, N.M. Shichova
Institute for Biology of Inland Waters RAS, 152742, Borok, Russia
The basic rules of the theory of errors are shortly stated. The main sources of errors originating at a quantitative estimation of a freshwater zoobenthos also at the analysis of growth and age of fishes are shown.